This week's installment of all things paleontological gives you a group of dinosaurs that prior to a large scale analysis remained something of the problem children of dinosaurian paleontology.
Some in depth information is included, but with luck it will be presented in an enjoyable manner!
In paleontology, mathematics is the paleontologist's Swiss army knife--from studying morphological change (how given traits shift and mold over vast stretches of time) to ancient ecological dynamics, without mathematics to guide our research and conclusions, paleontology would be largely knowledgeable guesswork.
One such mathematical tool, mentioned several times prior, is cladistic analysis.
The classic tree of life, with its uniform shape (ever leading towards what is of course the pinnacle evolution--mankind itself) with ever advancing improvement is drastically outdated. Rather, life as we know it can be described as analogous to a bush--ever branching, no particular direction or purpose past the essentials: individual reproduction and survival. And over time, with thousands of interactions between organisms and their environment, they change; through natural selection (both by environment stresses and reproductive benefits, such as the peacock's tail) isolation, and genetic drift, new populations and species arise from the old. No species is superior as a whole, just better suited for a given set of conditions. And if there's one thing we know about conditions, it's that they change constantly.

Cladistic analysis is the paleontologist's method of sorting out these ever branching evolutionary pathways, through labeling these morphological characters and using computer programs to sort them out and determine relationships therein. These trees are then mapped out in a visual diagram called a cladogram.
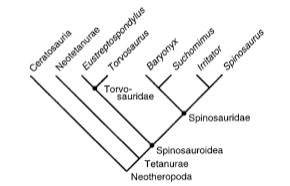
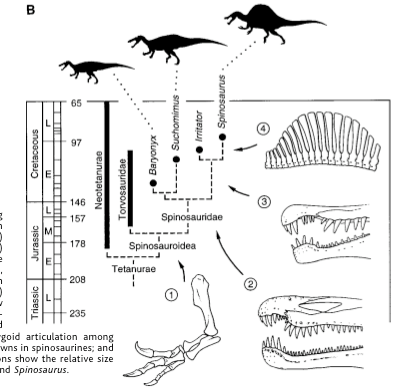
From the base of this branching structure, you have distantly related organisms to those in the top--each branch on this main line represents an evolutionary divergence, a shifting of structures (for example, placental mammals and marsupials both represent this kind of divergence--we both share a common ancestor that went in differing evolutionary pathways).
To construct a cladogram, a series of potential characters are listed under what is generally a binary code, like so (though this is just a single example!):
Baryonichae:
Dentary tooth count: 15 (0) 30 (1).
This suggests that for the Baryonichae (a subdivision of the prior mentioned Spinosauridae) having 30 teeth in the dentary, or lower jaw, is one of the states that helps unite the group--in which case a paleontologist running a cladistic analysis would place a '1' into the character matrix. Whereas if the creature in question had 15 teeth, he would place a zero for that particular species!
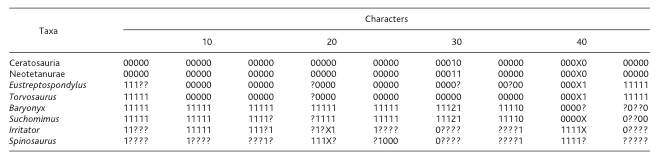
This particular matrix is somewhat limited due to the very fragmentary material of spinosaurids (only 45 characters listed)--some matrices have up to 1000 characters for dozens upon dozens or even hundreds of species!

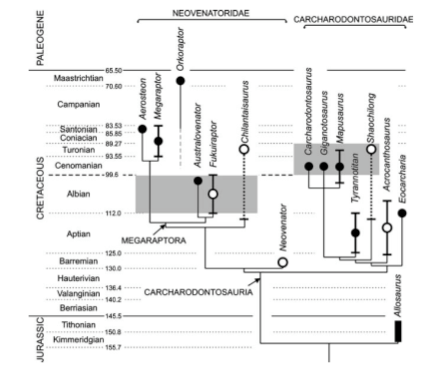
This then brings us to the primary subject of today's diary--the Neovenatoridae.
The Neovenatoridae is comprised of seven theropod dinosaurs spanning the globe (the British Neoventator, South American Megaraptor, Orkoraptor, and Aerosteon, Japanese Fukuiraptor, Chinese Chilantaisaurus, and Australian Australovenator) that had continuously perplexed paleontologists for years, continuously changing their placement within dinosaurian evolution to no truly stable and comfortable place. However, a recent analysis reveals in great detail their true nature, revealing fantastic biogeographic implications!

Part of the reason that it took such a long time to reveal their evolutionary relationships is their bizarre mix of characters--they all share long, gracile legs and elongate arms, a trait generally prescribed to the Coelurosauria, the group of dinosaurs containing both the tyrannosaurs and modern day birds. Still too they contain many features characteristic of Carcharodontosaurs (like the prior diaried Carcharodontosaurus) in the vertebral column amongst other regions.
Moreover, similar to dromaeosaurs like the famous Velociraptor, neovenatorids had a massively enlarged second digit toe class ('pedal ungual II', though relative to their body size not quite as extensive), causing several misidentifications in the past (an especially enigmatic member of the newly formed clade called Megaraptor has been labeled a spinosaurid, a dromaeosaur, and an allosaur in past analyses).

The analysis reveals that the Neovenatoridae is what we call the 'sister clade' to the Carcharodontosauridae (like one of my favorite dinosaurs, Carcharodontosaurus)--they both branched off in separate evolutionary directions from a distant common ancestor, the Neovinatorids developing a cursorial structure allowing good grasping and running power, while the Carcharodontosaurids opted for great size and bulk. These two groups together form a broader clade called the Carcharodontosauria.
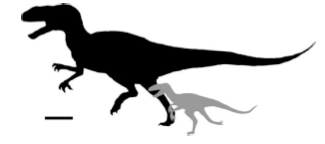
Adult sizes of these seven dinosaurs vary greatly, from a femur length of 50.7 centimeters to 119 centimeters. For just seven taxa, that's a pretty remarkable size difference.
Biogeographically (and temporally, in time), the relationships of these dinosaurs showed several significant things: one, the vicariance effect (evolutionary divergence caused by geographic isolation) though existent, was not quite as intensive a factor in dinosaurian evolution as previously thought, given the global distribution of the animals (rather local extinctions, which opened up new available ecological niches and evolutionary innovation, combined with vicariance helped craft the dinosaurs into the forms they took). Secondly, it was previously suggested that in the southern hemisphere the allosaurids (of which the carcharodontosaurs were an advanced, later form) became extinct and that only the previously diaried abelisaurs remained. With the very late cretaceous Orkoraptor, we know that this is not the case!
But that's part of the beauty of paleontology. It combines adventure and exploration (both through the physical medium of fieldwork and mental exploration in the labs and museums across the world) with the tools and rigorous testing and questioning essential for science into an always shifting and enhanced understanding. Unlike creationists (who come to their conclusions preemptively and pick and choose data to match that conclusion), scientists largely don't claim to have the whole truth--rather, they collect data, interpret the available data, test it, and collect more, all the while enhancing our understanding of ancient worlds past.
Paul Sereno described paleontology best as "adventure with a purpose," and it's that adventure and unending curiosity that drives me in my particular field.

Enjoy, and feel free to rec away!
